Action potential
2008/9 Schools Wikipedia Selection. Related subjects: Biology

In neurophysiology, an action potential (also known as a nerve impulse or spike) is a pulse-like wave of voltage that travels along several types of cell membranes. The best-understood example is generated on the membrane of the axon of a neuron, but also appears in other types of excitable cells, such as cardiac muscle cells, and even plant cells. The resting voltage across the axonal membrane is typically −70 millivolts (mV), with the inside being more negative than the outside. As an action potential passes through a point, this voltage rises to roughly +40 mV in one millisecond, then returns to −70 mV. The action potential moves rapidly down the axon, with a conduction velocity as high as 100 meters/second (224 miles per hour). Because they are able to transmit information so fast, the flow of action potentials is a very efficient form of data transmission, considering that each neuron the signal passes through can be up to a meter in length.
An action potential is provoked on a patch of membrane when the membrane is depolarized sufficiently, i.e., when the voltage of the cell's interior relative to the cell's exterior is raised above a threshold. Such a depolarization opens voltage-sensitive channels, which allows current into the axon, further depolarizing the membrane. This will cause the membrane to "fire", initiating a positive feedback loop that suddenly and rapidly causes the voltage inside the axon to become more positive. After this rapid rise, the membrane voltage is restored to its resting value by a combination of effects: the channels responsible for the initial inward current are inactivated, while the raised voltage opens other voltage-sensitive channels that allow a compensating outward current. Because of the positive feedback, an action potential is all-or-none; there are no partial action potentials. In neurons, a typical action potential lasts for just a few thousandths of a second at any given point along their length. The passage of an action potential can leave the ion channels in a non-equilibrium state, making them more difficult to open, and thus inhibiting another action potential at the same spot: such an axon is said to be refractory.
The principal ions involved in an action potential are sodium and potassium cations; sodium ions enter the cell, and potassium ions leave, restoring equilibrium. Relatively few ions need to cross the membrane for the membrane voltage to change drastically. The ions exchanged during an action potential, therefore, make a negligible change in the interior and exterior ionic concentrations. The few ions that do cross are pumped out again by the continual action of the sodium–potassium pump, which, with other ion transporters, maintains the normal ratio of ion concentrations across the membrane. Calcium cations and chloride anions are involved in a few types of action potentials, such as the cardiac action potential and the action potential in the single-celled alga Acetabularia, respectively.
The action potential "travels" along the axon without fading out because the signal is regenerated at each patch of membrane. This happens because an action potential at one patch raises the voltage at nearby patches, depolarizing them and provoking a new action potential there. In unmyelinated neurons, the patches are adjacent, but in myelinated neurons, the action potential "hops" between distant patches, making the process both faster and more efficient. The axons of neurons generally branch, and an action potential often travels along both forks from a branch point. The action potential stops at the end of these branches, but usually causes the secretion of neurotransmitters at the synapses that are found there. These neurotransmitters bind to receptors on adjacent cells. These receptors are themselves ion channels, although—in contrast to the axonal channels—they are generally opened by the presence of a neurotransmitter, rather than by changes in voltage. The opening of these receptor channels can help to depolarize the membrane of the new cell (an excitatory channel) or work against its depolarization (an inhibitory channel). If these depolarizations are sufficiently strong, they can provoke another action potential in the new cell.
Biophysical and cellular context
Ions and the forces driving their motion

Electrical signals within biological organisms are generally by ions, which may be either positively charged cations or negatively charged anions. The most important cations for the action potential are sodium (Na+) and potassium (K+), which are both monovalent cations that carry a single positive charge. Action potentials can also involve calcium (Ca2+), which is a divalent cation that carries a double positive charge. The chloride anion (Cl−) plays a major role in the action potentials of some algae, but plays a negligible role in the action potentials of most animals.
Ions cross the cell membrane under two influences: diffusion and electric fields. Diffusion allows net flow of ions from regions where the ions are highly concentrated into regions of low concentration. Ions also move in response to an electric field. By definition, the integral of the electric field across a patch of membrane equals the voltage Vm across that patch. Likewise by definition, the flows of different ions through that patch are the ionic currents at that patch; the total current is the sum of all the individual ionic currents. Using these definitions of voltage and current, such a membrane patch can be modeled by an equivalent electronic circuit. In particular, for each type of ion the patch will have a capacitance C and a conductance g; according to Ohm's law, the current I of each ion type is related to the transmembrane voltage Vm by the equation I = g Vm. For a given set of ionic conductances, there is an equilibrium voltage E at which the total current across the membrane is zero; the natural flow of ions generally causes the membrane voltage Vm to approach E.

Cell membrane
Because the membrane surrounding cells is nearly impermeable to ions, cells have evolved systems for transporting ions across the membrane. These systems can be divided into two classes: pores ("channels") that allow passive transport of ions, and ion pumps that use adenosine triphosphate for active transport of ions. The ion pumps tend to work continuously, as long as there are ions to be pumped. By contrast, the ion channels open and close in response to signals from their environment. The two classes play complementary roles; the ion pumps generate the differences in ion concentrations across the membrane, which the ion channels exploit to carry out electrical signaling. As an analogy, ion pumps play the role of the battery that allows a radio circuit (the ion channels) to transmit a signal.

Ion channels
Ion channels are integral membrane proteins through which ions can cross the membrane. Most channels are specific for one ion; whereas that ion passes through relatively quickly, other similar ions pass through very infrequently. For example, although potassium and sodium ions have the same charge and differ only slightly in their radius, potassium channels allow few sodium ions through, and vice versa. The pore through which the ion passes is typically so small that ions must pass through it alone and single-file. Channels are either fully open or fully closed. When the channel is open, ions flow through it by passive transport, i.e., at a rate determined by the membrane voltage Vm and concentration difference across the membrane. The action potential is a manifestation of different ion channels opening and closing at different times.

A channel may have several different states (corresponding to different conformations of the protein), but each such state is either open or closed. In general, closed states correspond either to a contraction of the pore—making it impassable to the ion—or to a separate part of the protein stoppering the pore. For example, the voltage-dependent sodium channel undergoes inactivation, in which a portion of the protein swings into the pore, sealing it. This inactivation shuts off the sodium current and plays a critical role in the action potential.
Ion channels can be classified by how they respond to their environment. For example, the ion channels involved in the action potential are voltage-sensitive channels; they open and close in response to the voltage across the membrane. Ligand-gated channels form another important class; these ion channels open and close in response to the binding of a ligand molecule, such as a neurotransmitter. Other ion channels open and close with mechanical forces. Still other ion channels—such as those of sensory neurons—open and close in response to other stimuli, such as light, temperature or pressure.
Ion pumps
The ionic currents of the action potential flow in response to concentration differences of the ions across the cell membrane. These concentration differences are established by ion transporters, which are integral membrane proteins that carry out active transport, i.e., use cellular energy (ATP) to "pump" the ions against their concentration gradient. Such ion pumps take in ions from one side of the membrane (decreasing its concentration there) and release them on the other side (increasing its concentration there). The ion pump most relevant to the action potential is the sodium–potassium pump, which transports three sodium ions out of the cell and two potassium ions in. Consequently, the concentration of potassium ions K+ inside the neuron is roughly 20-fold larger than the outside concentration, whereas the sodium concentration outside is roughly ninefold larger than inside. Similarly, other ions have different concentrations inside and outside the neuron, such as calcium, chloride and magnesium.
Ion pumps influence the action potential only by establishing the relative ratio of intracellular and extracellular ion concentrations. The action potential mainly involves the opening and closing of ion channels, not ion pumps. If the ion pumps are turned off by removing their energy source, or by adding an inhibitor such as ouabain, the axon can still fire hundreds of thousands of action potentials before their amplitudes begin to decay significantly. In particular, ion pumps play no significant role in the repolarization of the membrane after an action potential.
Resting potential
Each type of ion has a reversal potential E—also called its equilibrium voltage or equilibrium potential—at which the net current of that ion across the membrane is zero; the ionic current due to the electric field exactly cancels the current due to the differences in concentration across the membrane. That equilibrium voltage is given by the Nernst equation
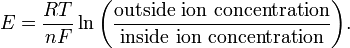
The constants in this equation are the charge valence n of the ion (e.g., +1 for K+, +2 for Ca2+ and −1 for Cl−), the temperature T (in Kelvins), the molar gas constant R, and the Faraday F, which is the total charge of a mole of electrons. For illustration, at a typical physiological ratio of concentrations, the potassium equilibrium voltage EK is −75 mV, whereas the sodium equilibrium voltage ENa is +55mV. Since these disagree, there is no voltage at which the currents of potassium and sodium ions are both zero.
However, there is a voltage Em at which the net current of all ions across the membrane is zero; this voltage is given by the Goldman equation
![E_{m} = \frac{RT}{F} \ln{ \left( \frac{ P_{\mathrm{K}}[\mathrm{K}^{+}]_\mathrm{out} + P_{\mathrm{Na}}[\mathrm{Na}^{+}]_\mathrm{out} + P_{\mathrm{Cl}^{-}}[\mathrm{Cl}^{-}]_\mathrm{in}}{ P_{\mathrm{K}}[\mathrm{K}^{+}]_\mathrm{in} + P_{\mathrm{Na}}[\mathrm{Na}^{+}]_\mathrm{in} + P_{\mathrm{Cl}}[\mathrm{Cl}^{-}]_\mathrm{out}} \right) }](../../images/782/78240.png)
for the three monovalent ions most important to action potentials: potassium (K+), sodium (Na+), and chloride (Cl−). Being an anion, the chloride terms are treated differently than the cation terms; the inside concentration is in the numerator, and the outside concentration is in the denominator, which is reversed from the cation terms. Pi stands for the permeability of the ion type i. If calcium ions are also considered, the formula for the equilibrium voltage becomes more complicated.
The membrane voltage Vm need not equal its equilibrium value Em. However, since Vm can change drastically when only a few ions cross the membrane, Vm tracks Em closely, so that the two are effectively equivalent. In a typical action potential, where Vm changes by roughly 100 mV, the ionic concentrations inside the axon change only by roughly 1 part in 10 million; hence, hundreds of thousands of action potentials can be fired before the ion pumps are needed to restore the standard ratio of ionic concentrations.
The equilibrium voltage Em under normal, unstimulated conditions is called the resting potential Vrest, typically −70 mV. (The word "potential" or "potential difference" is sometimes a synonym for voltage.) Under those conditions, the membrane is much more permeable to potassium than to any other ion; thus, consistent with the Goldman equation, the resting potential is close to the potassium equilibrium potential EK. In the middle of the action potential, however, the sodium permeability dominates, so that Em is +45 mV, close to the sodium equilibrium voltage ENa.

Anatomy of a neuron
Several types of cells support an action potential, such as plant cells, muscle cells, and the specialized cells of the heart (in which occurs the cardiac action potential). However, the main excitable cell is the neuron, which also has the simplest mechanism for the action potential.
Most neurons have numerous branched tendrils called neurites, which are divided into two main types, dendrites and axons. Most neurons have only one axon but numerous dendrites; the beginning of the axon is called the axon hillock. Action potentials almost always begin at the axon hillock, and travel down the axon; it is very rare for an action potential to occur in the dendrites. A typical axon has a few branch points, forming a tree-like shape; the tips of this tree (the axonal termini) are generally called the synaptic knobs. These knobs are usually adjacent to the dendrites of another neuron or, more generally, to another excitable cell; the contact between them is called the synapse.
| Neuron |
|---|

In some animals (mostly vertebrates), segments of the axon are sheathed in myelin, which generally increases the conduction velocity at which action potentials travel down the axon. Myelin is composed of Schwann cells that wrap themselves multiple times around the axonal segment, forming a thick fatty layer that prevents ions from entering or escaping the axon. Ions can flow into and out of the axon only at the nodes of Ranvier, which are the gaps between the Schwann cells, the "chinks" in the myelin armor. Therefore, the action potential "hops" from one node of Ranvier to the next (the process of saltatory conduction); it does not move continuously down the axon, as it does in unmyelinated axons.
Phases
The course of the action potential is determined by two coupled effects. First, voltage-sensitive ion channels open and close in response to changes in the membrane voltage Vm, thus changing the membrane's permeability to those ions. However, by the Goldman equation, changes in the ionic permeabilities causes changes in the equilibrium potential Em, and, thus, the membrane voltage Vm. This two-way interaction between Vm and the ion channels sets up the possibility for positive feedback, which is a key part of the rising phase of the action potential. A complicating factor is that a single ion channel may have multiple internal "gates" that respond to changes in Vm in opposite ways, or at different rates. For example, although raising Vm opens most gates in the voltage-sensitive sodium channel, it also closes the channel's "inactivation gate", albeit more slowly. Hence, when Vm is raised suddenly, the sodium channels open initially, but then close due to the slower inactivation.
The course of the action potential can be divided into four parts: the rising phase, the falling phase, the undershoot phase, and the refractory period. The initial membrane permeability to potassium is low, but much higher than that of other ions, making the resting potential close to EK. A sufficiently strong depolarization (increase in Vm) causes the voltage-sensitive sodium channels to open; the increasing permeability to sodium drives Vm closer to the sodium equilibrium voltage ENa≈ +55 mV. The increasing voltage in turn causes even more sodium channels to open, which pushes Vm still further towards ENa. This positive feedback continues until the sodium channels are fully open and Vm is close to ENa. This is the rising phase. At this point, the sodium channels begin to inactivate, lowering the membrane's permeability to sodium and driving Vm back down toward the original resting potential. Meanwhile, the potassium channels open more fully; the increased permeability to potassium likewise helps to drive the membrane voltage back down towards EK, the potassium equilibrium voltage. This is the falling phase. The potassium conductance remains unusually high, causing the membrane voltage to dip below even the resting potential; this is the undershoot phase. Finally, the time during which a subsequent action potential is impossible or difficult to fire is called the refractory period, which may overlap with the other phases.
The voltages and currents of the action potential in all of its phases were modeled accurately by Alan Lloyd Hodgkin and Andrew Huxley in 1952, for which they were awarded the Nobel Prize in Physiology or Medicine in 1963. However, their model considers only two types of voltage-sensitive ion channels, and makes several assumptions about them, e.g., that their internal gates open and close independently of one another. In reality, there are many types of ion channels, and they do not always open and close independently.
Stimulation and rising phase
A typical action potential begins at the axon hillock with a sufficiently strong depolarization, e.g., a stimulus that increases Vm. This depolarization is often caused by the injection of extra sodium cations into the cell; these cations can come from a wide variety of sources, such as chemical synapses, sensory neurons or pacemaker potentials.
The depolarization opens both the sodium and potassium channels in the membrane, allowing the ions to flow into and out of the axon, respectively. If the depolarization is small (say, increasing Vm from −70 mV to −60 mV), the outward potassium current overwhelms the inward sodium current and the membrane repolarizes back to its normal resting potential around −70 mV. The "failed initiations" shown in Figure 1 illustrate this response. However, if the depolarization is large enough, the inward sodium current increases more than the outward potassium current and a runaway condition ( positive feedback) results: the more inward current there is, the more Vm increases, which in turn further increases the inward current. The sharp rise in Vm and sodium permeability correspond to the rising phase of the action potential.
The critical threshold voltage for this runaway condition is usually around −45 mV, but it depends on the recent activity of the axon. A membrane that has just fired an action potential cannot fire another one immediately, since the ion channels have not returned to their usual state. The period during which no new action potential can be fired is called the absolute refractory period. At longer times, after some but not all of the ion channels have recovered, the axon can be stimulated to produce another action potential, but only with a much stronger depolarization, e.g., −30 mV. The period during which action potentials are unusually difficult to provoke is called the relative refractory period.
Peak and falling phase
The positive feedback of the rising phase slows and comes to a halt as the sodium ion channels become maximally open. At the peak of the action potential, the sodium permeability is maximized and the membrane voltage Vm is nearly equal to the sodium equilibrium voltage ENa. However, the same raised voltage that opened the sodium channels initially also slowly shuts them off, by stoppering their pores; the sodium channels become inactivated. This lowers the membrane's permeability to sodium, driving the membrane voltage back down. At the same time, the raised voltage opens voltage-sensitive potassium channels; the increase in the membrane's potassium permeability drives Vm towards EK. Combined, these changes in sodium and potassium permeability cause Vm to drop quickly, repolarizing the membrane and producing the "falling phase" of the action potential.
Hyperpolarization ("undershoot")
The raised voltage opened many more potassium channels than usual, and these do not close right away when the membrane returns to its normal resting voltage. The potassium permeability of the membrane is transiently unusually high, driving the membrane voltage Vm even closer to the potassium equilibrium voltage EK. Hence, there is an undershoot, a hyperpolarization in technical language, that persists until the membrane potassium permeability returns to its usual value.
Refractory period
The opening and closing of the sodium and potassium channels during an action potential may leave some of them in a "refractory" state, in which they are unable to open again until they have recovered. In the absolute refractory period, so many ion channels are refractory that no new action potential can be fired. Significant recovery (de-inactivation) requires that the membrane potential remain hyperpolarized for a certain length of time. In the relative refractory period, enough channels have recovered that an action potential can be provoked, but only with a stimulus much stronger than usual. These refractory periods ensure that the action potential travels in only one direction along the axon.
Initiation, propagation and termination
A typical action potential is initiated at the axon hillock when the membrane is depolarized sufficiently, i.e., when its voltage is increased sufficiently. As the membrane voltage is increased, both the sodium and potassium ion channels begin to open up, increasing both the inward sodium current and the balancing outward potassium current. For small voltage increases, the potassium current triumphs over the sodium current and the voltage returns to its normal resting value, typically −70 mV. However, if the voltage increases past a critical threshold, typically 15 mV higher than the resting value, the sodium current dominates and a runaway condition results; the cell "fires", producing an action potential. Once started, the action potential propagates down the axon without diminishing; the inwards current of an action potential at one patch of membrane depolarizes nearby membrane patches, sparking another action potential there. In effect, the action potential is created afresh at each patch of membrane; its energy derives from the local differences in ionic concentrations, not from the depolarization that triggered it. The axon may branch along its length, and there the inward current may not quite suffice to trigger a new action potential in one or both of its branches; the action potential may stop. Action potentials that do reach the ends of the axon generally cause the release of a neurotransmitter into the synapse, which may combine with other inputs to provoke a new action potential in the post-synaptic neuron or muscle cell.
Initiation
Before considering the propagation of action potentials along axons and their termination at the synaptic knobs, it is helpful to consider the methods by which action potentials can be initiated at the axon hillock. The basic requirement is that the membrane voltage at the hillock be raised above the threshold for firing; there are several ways in which this depolarization can occur.

Neurotransmission
Action potentials are most commonly initiated by excitatory postsynaptic potentials from a presynaptic neuron. Typically, neurotransmitter molecules are released by the presynaptic neuron bound to receptors on the postsynaptic cell. This binding opens various types of ion channels, changing the local permeability of the cell membrane and thereby altering the membrane potential. If the binding increases the voltage (depolarizes the membrane), the synapse is excitatory; if the binding decreases the voltage (hyperpolarizes the membrane), it is inhibitory. Whether the voltage is decreased or increased, the change propagates passively to nearby regions of the membrane, as described by the cable equation and its refinements; typically, the voltage stimulus decays exponentially with the distance from the synapse and with time from the binding of the neurotransmitter. Some fraction of an excitatory voltage may reach the axon hillock and may (in rare cases) depolarize the membrane enough to provoke a new action potential. More typically, the excitatory potentials from several synapses must work together at nearly the same time to provoke a new action potential. Their joint efforts can be thwarted, however, by the counter-acting inhibitory postsynaptic potentials.
Neurotransmission can also occur through electrical synapses. Due to the direct connection between the excitable cells in such cases, an action potential can well be transmitted directly from one cell to the next. Rectifying channels ensure that action potentials only move in one direction through an electrical synapse.
Sensory neurons
In sensory neurons, an external signal such as pressure, temperature, light, or sound is coupled with the opening and closing of ion channels, which in turn alter the ionic permeabilities of the membrane and its voltage. These voltage changes can again be excitatory (depolarizing) or inhibitory (hyperpolarizing) and, in some sensory neurons, their combined effects can depolarize the axon hillock enough to provoke action potentials. Examples in humans include the olfactory receptor neuron and Meissner's corpuscle, which are critical for the sense of smell and touch, respectively. However, not all sensory neurons convert their external signals into action potentials; some do not even have an axon! Instead, they may convert the signal into the release of a neurotransmitter, or into continuous graded potentials, either of which may stimulate subsequent neuron(s) into firing an action potential. For illustration, in the human ear, hair cells convert the incoming sound into the opening and closing of mechanically gated ion channels, which may cause neurotransmitter molecules to be released. Similarly, in the human retina, the initial photoreceptor cells and the next two layers of cells ( bipolar cells and amacrine cells) do not produce action potentials; only the third layer, the ganglion cells, produce action potentials, which then travel up the optic nerve.
Pacemaker potentials

In the cases of neurotransmission and sensory neurons, action potentials result from an external stimulus. However, some excitable cells require no such stimulus to fire: they spontaneously depolarize their axon hillock and fire action potentials at a regular rate, like an internal clock. The voltage traces of such cells are known as pacemaker potentials. The cardiac pacemaker cells of the sinoatrial node in the heart provide a good example. Although such pacemaker potentials have a natural rhythm, it can be adjusted by external stimuli; for instance, heart rate can be altered by pharmaceuticals as well as signals from the sympathetic and parasympathetic nerves. The external stimuli do not cause the cell's repetitive firing, but merely alter its timing. In some cases, the regulation of frequency can be more complex, leading to patterns of action potentials, such as bursting.
Propagation
The action potential propagates as a wave along the axon. The currents flowing inwards at a point on the axon during an action potential spread out along the axon, and depolarize the adjacent sections of its membrane. If sufficiently strong, this depolarization provokes a similar action potential at the neighboring membrane patches. This basic mechanism was demonstrated by Alan Lloyd Hodgkin in 1937. After crushing or cooling nerve segments and thus blocking the action potentials, he showed that an action potential arriving on one side of the block could provoke another action potential on the other, provided that the blocked segment was sufficiently short.
Once an action potential has occurred at a patch of membrane, the membrane patch needs time to recover before it can fire again. At the molecular level, this absolute refractory period corresponds to the time required for its ion channels to return to their normal open or closed states. Although it limits the frequency of firing, the absolute refractory period ensures that the action potential moves in only one direction along an axon. The currents flowing in due to an action potential spread out in both directions along the axon. However, only the unfired part of the axon can respond with an action potential; the part that has just fired is unresponsive until the action potential is safely out of range and cannot restimulate that part. In the usual orthodromic conduction, the action potential propagates from the axon hillock towards the synaptic knobs (the axonal termini); propagation in the opposite direction—known as antidromic conduction—is very rare. However, if a laboratory axon is stimulated in its middle, both halves of the axon are "fresh", i.e., unfired; then two action potentials will be generated, one traveling towards the axon hillock and the other traveling towards the synaptic knobs.

Myelin and saltatory conduction
The axons of some neurons are ensheathed in myelin, a fatty (ie, lipid-rich) insulating material that increases the speed and energy efficiency of action potential conduction. Axons are myelinated by specialized cells, Schwann cells and oligodendrocytes, that wrap themselves multiple times around segments of axon. The gaps between these segments are known as the nodes of Ranvier.
Myelin prevents ions from entering or leaving the axon along myelinated segments. Myelination is found mainly in vertebrates, but an analogous system has been discovered in a few invertebrates, such as some species of shrimp. As a general rule, myelination increases the conduction velocity of action potentials and makes them more energy-efficient. However, not all neurons in vertebrates are myelinated. Whether saltatory or not, the mean conduction velocity of an action potential ranges from 1 m/s to over 100 m/s, and generally increases with axonal diameter.
Action potentials cannot propagate through the myelinated segments of the axon, since no ions can flow across the membrane there. Instead, the ionic current from an action potential at one node of Ranvier provokes another action potential at the next node; this "hopping" of the action potential from node to node is known as saltatory conduction. Although the mechanism of saltatory conduction was suggested in 1925 by Ralph Lillie, the first experimental evidence for saltatory conduction came from Ichiji Tasaki and Taiji Takeuchi and from Alan Hodgkin and Robert Stämpfli. By contrast, in unmyelinated axons, the action potential provokes another in the membrane immediately adjacent, and moves continuously down the axon like a wave.

Myelin has two important advantages: fast conduction speed and energy efficiency. For axons larger than a minimum diameter (roughly 1 micron), myelination increases the conduction velocity of an action potential, typically tenfold. Conversely, for a given conduction velocity, myelinated fibers are smaller than their unmyelinated counterparts. For example, action potentials move at roughly the same speed (25 m/s) in a myelinated frog axon and an unmyelinated squid giant axon, but the frog axon has a roughly 30-fold smaller diameter and 100-fold smaller cross-sectional area. Also, since the ionic currents are confined to the nodes of Ranvier, far fewer ions "leak" across the membrane, saving metabolic energy. This saving is a significant selective advantage, since the human nervous system uses approximately 20% of the body's metabolic energy.
The length of axons' myelinated segments is important to the success of saltatory conduction. They should be as long as possible to maximize the speed of conduction, but not so long that the arriving signal is too weak to provoke an action potential at the next node of Ranvier. In nature, myelinated segments are generally long enough for the passively propagated signal to travel for at least two nodes while retaining enough amplitude to fire an action potential at the second or third node. Thus, the safety factor of saltatory conduction is high, allowing transmission to bypass nodes in case of injury. However, action potentials may end prematurely in certain places where the safety factor is low, even in unmyelinated neurons; a common example is the branch point of an axon, where it divides into two axons.
Some diseases degrade myelin and impair saltatory conduction, reducing the conduction velocity of action potentials. The most well-known of these is multiple sclerosis, in which the breakdown of myelin impairs coordinated movement.
Cable theory

The flow of currents within an axon can be described quantitatively by cable theory and its elaborations, such as the compartmental model. Cable theory was developed in 1855 by Lord Kelvin to model the transatlantic telegraph cable and was shown to be relevant to neurons by Hodgkin and Rushton in 1946. In simple cable theory, the neuron is treated as an electrically passive, perfectly cylindrical transmission cable, which can be described by a partial differential equation
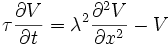
where V(x, t) is the voltage across the membrane at a time t and a position x along the length of the neuron, and where λ and τ are the characteristic length and time scales on which those voltages decay in response to a stimulus. Referring to the circuit diagram above, these scales can be determined from the resistances and capacitances per unit length
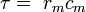
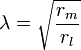
These time- and length-scales can be used to understand the dependence of the conduction velocity on the diameter of the neuron in unmyelinated fibers. For example, the time-scale τ increases with both the membrane resistance rm and capacitance cm. As the capacitance increases, more charge must be transferred to produce a given transmembrane voltage (by the equation Q=CV); as the resistance increases, less charge is transferred per unit time, making the equilibration slower. Similarly, if the internal resistance per unit length ri is lower in one axon than in another (e.g., because the radius of the former is larger), the spatial decay length λ becomes longer and the conduction velocity of an action potential should increase. If the transmembrane resistance rm is increased, that lowers the average "leakage" current across the membrane, likewise causing λ to become longer, increasing the conduction velocity.
Termination
Chemical synapses
Action potentials that reach the synaptic knobs generally cause a neurotransmitter to be released into the synaptic cleft. Neurotransmitters are small molecules that may open ion channels in the postsynaptic cell; most axons have the same neurotransmitter at all of their termini. The arrival of the action potential opens voltage-sensitive calcium channels in the presynaptic membrane; the influx of calcium causes vesicles filled with neurotransmitter to migrate to the cell's surface and release their contents into the synaptic cleft. This complex process is inhib